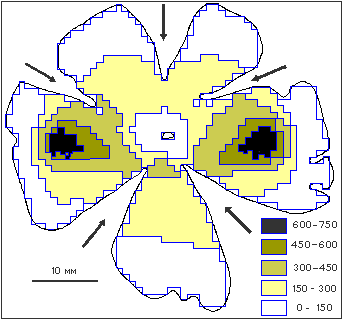
Всадники Перна. МатериалыДельфины -> Что видит и слышит дельфинА. М. Масс, А. Я. СупинЛет двадцать тому назад дельфины были в большой моде. Не было недостатка в фантастических спекуляциях во всем, что касалось этих животных. Со временем мода прошла, и спекуляции заслуженно забыты. А что же осталось? То, что привлекало ученых с самого начала. Дельфины целиком приспособлены к жизни в воде, все системы их организма (органы чувств, дыхание, кровообращение и многое другое) работают в совершенно иных условиях, чем аналогичные системы, например человека. Среди всех систем организма дельфина, нас привлекли зрительная и слуховая. Дело в том, что, проводя всю жизнь в воде, дельфин все же вынужден постоянно всплывать на поверхность для дыхания. Поэтому его зрение должно быть достаточно хорошим и под водой, и в воздухе, а это непросто, поскольку оптические свойства воздуха и воды совершенно различны. С другой стороны, каким бы хорошим ни было зрение дельфина, его возможности ограничены из-за невысокой прозрачности воды. Поэтому основные сведения об окружающей обстановке дельфин получает с помощью слуха. При этом он использует активную локацию: слушает эхо, возникающее при отражении издаваемых им звуков от окружающих предметов. Эхо дает ему точные сведения не только о положении предметов, но и об их величине, форме, материале. Иными словами, слух позволяет дельфину воспринимать окружающий мир не хуже или даже лучше, чем зрение. Что может увидеть дельфинОбычно, чтобы это определить, проводят эксперименты с условными рефлексами [1]. Но не всегда есть условия для выполнения таких исследований на дельфинах, а тем более на их более крупных сородичах - китах. Задачу помогает решить морфологическое изучение глаза, в частности строение сетчатки, которое связано с особенностями зрения. Фоторецепторы и нервные клетки распределены в сетчатке неравномерно. Например, у человека и приматов существует участок около 0.5 мм2, называемый желтым пятном. В этой области плотность рецепторов максимальна. Именно эта область имеет наибольшую остроту зрения и представлена в высших отделах мозга наиболее подробно, благодаря чему мы видим предмет детально. Подобные области наилучшего видения (ОНВ) в той или иной форме существуют у всех животных. Размер, форма, расположение ОНВ зависят от образа жизни животного и положения глаз на голове. У человека и животных с фронтальным расположением глаз (у приматов, хищных) эта область представлена в виде пятнышка в центральной части сетчатки, у животных с глазами, расположенными по бокам головы (у грызунов, копытных), эта зона выглядит в виде горизонтально вытянутой полоски. До недавнего времени считалось, что у китов и дельфинов ОНВ отсутствует, однако по нашим данным ОНВ есть и у морских млекопитающих, в том числе у дельфинов и китов. Острота зрения животных может быть рассчитана по плотности ганглиозных клеток сетчатки [2]. Эти нервные клетки расположены в сетчатке глаза и производят предварительную обработку информации, приходящей от светочувствительных рецепторов. Роль ганглиозных клеток в процессах зрения очень велика, и, в частности, способность сетчатки глаза различать мелкие детали изображения зависит от того, насколько плотно расположены в ней эти клетки: чем их больше на единицу площади, тем более мелкие детали изображения могут различаться. Подсчитывая плотность, т. е. количество ганглиозных клеток на единицу площади, можно оценить остроту зрения в той или иной части поля зрения, в том числе и найти ОНВ. Объектом нашего исследования стал хорошо известный и исследователям, и зрителям океанариумов черноморский дельфин афалина ( Tursiоps truncatus). Данные о его остроте зрения особенно важны как для проведения экспериментальных исследований, так и для создания необходимых условий для содержания этих уникальных животных в океанариумах. Более 10 лет мы собирали морфологический материал от погибших (случайно или в результате болезни) дельфинов. Это позволило получить достаточное количество препаратов сетчатки, чтобы провести полноценное исследование [3]. Если сетчатку целиком извлечь из глазного бокала, а затем наклеить на предметное стекло и окрасить специальным красителем, то получается препарат, который можно обследовать под обычным световым микроскопом, подсчитать количество и установить плотность расположения ганглиозных клеток. Таким образом составлялась топографическая карта сетчатки, показывающая, как распределены в ней ганглиозные клетки (рис.1).
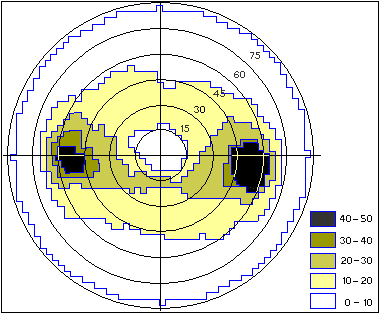
Плотность клеток (число клеток на 1 мм2) условно обозначена штриховкой в соответствии со шкалой. Слева - передний, справа - задний, сверху и снизу - верхний и нижний полюсы сетчатки соответственно. Стрелками показаны разрезы, сделанные для наклеивания сетчатки на плоское стекло Обнаружилась удивительная картина. В отличие от всех наземных млекопитающих, имеющих одну ОНВ, в сетчатке дельфина существуют две такие области. Расположены они на экваторе сетчатки: одна из них находится в задней части, т.е. в той, которая смотрит вперед (ведь оптическая система глаза переворачивает изображение); другая - в передней, которая смотрит назад. Эти области расположены довольно далеко от центра сетчатки, на расстоянии 15-17 мм. Плотность ганглиозных клеток оказалась примерно одинаковой в обеих зонах: около 670 клеток/мм2. На полученной карте (рис.1) представлена вся сетчатка, развернутая на плоскости. Для того чтобы наклеить сферическую сетчатку на плоское стекло, пришлось сделать на ней несколько радиальных надрезов. А чтобы оценить, как организовано поле зрения дельфина, полученная карта “сшивается” по линиям разреза с помощью специальной компьютерной программы. В результате получается непрерывная сферическая карта распределения ганглиозных клеток в полярных координатах (рис.2). Зная радиус сетчатки (14.5 мм), мы рассчитали, что максимальная плотность составляет 43 клетки/град2. Это значит, что среднее угловое расстояние между соседними клетками составляет примерно 9'. Острота зрения определяется именно этим угловым расстоянием. Значит, острота зрения дельфина в обеих ОНВ (и в передней, и в задней) - около 9'.
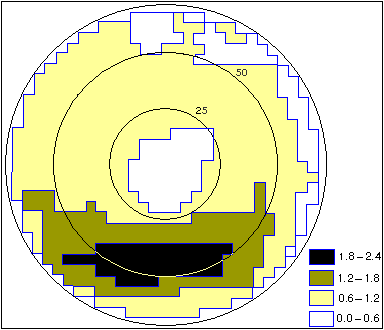
Плотность клеток (число клеток на 1 град2) условно обозначена штриховкой в соответствии со шкалой. Цифры у концентрических окружностей - расстояние от центра сетчатки (в угловых градусах) Эти расчеты были сделаны с учетом оптических свойств воды. А какова острота зрения дельфина в воздушной среде? По законам оптики, под водой изображение предмета на сетчатке примерно на треть больше, чем изображение того же предмета, расположенного на том же расстоянии, но в воздухе. Поэтому в воздушной среде способность сетчатки дельфина различать изображения немного хуже, чем в воде: острота зрения составляет 12'. Выявленная картина топографии сетчатки китообразных с двумя ОНВ совершенно необычна и не имеет аналогов ни у одного из наземных млекопитающих. Это особенно интересно, если учитывать, что дельфины и киты - вторичноводные животные (их предки жили когда-то на суше). Но для чего же дельфину две ОНВ вместо одной? Оптическая система, рассчитанная на работу в воздухе, дает плохое изображение под водой, и наоборот. Дельфины же хорошо видят и под водой, и на воздухе. Не исключено, что такая универсальность зрения дельфинов и связана с наличием в их глазе двух ОНВ: на одной из них оптика глаза обеспечивает хорошее изображение в воде, а на другой - преимущественно в воздухе. Это предположение подтверждается многими наблюдениями. Когда дельфин хочет что-то рассмотреть под водой, он обычно поворачивается к объекту боком (одним глазом), т.е. использует заднебоковую ОНВ. А чтобы увидеть предмет в воздухе, дельфин располагается к нему носом (смотрит двумя глазами), т.е. использует переднюю ОНВ. Интересно было узнать, характерно ли такое необычное строение глаза только для афалины или для других дельфинов тоже? К настоящему времени сведения о строении сетчатки получены для нескольких видов дельфинов [4-6]. У всех у них сетчатка глаза содержит две ОНВ. И острота зрения тоже более или менее сходная: 8-14'. Это хуже, чем у многих наземных животных: например, у человека и приматов острота зрения около 1', а у кошки 5-6'. Но для подводных условий, где прозрачность среды невелика, этого вполне достаточно. Это доказывают и наблюдения за животными, содержащимися в океанариумах: они прекрасно знают своих тренеров, могут совершать точные прицельные прыжки и без единого промаха доставать до предметов, помещенных на большой высоте. Исключение составил только амазонский дельфин ( Inia geoffrensis) - удивительное животное, обитающее не в морях, а в реках Южной Америки. Вода там очень мутная, почти непрозрачная, и глаза амазонского дельфина приспособились к зрению на очень коротком расстоянии (дальше все равно ничего не видно) и при низкой освещенности (мутная вода сильно поглощает свет). У этого дельфина только одна ОНВ, но не в центре сетчатки, как у наземных животных, а в нижней ее части - той, которая смотрит вверх (рис.3). Только там, в верхнем слое воды, достаточно света, чтобы хоть что-нибудь разглядеть. Соответственно и острота зрения у амазонского дельфина очень низкая: 45' [7]. Но большая острота зрения и не нужна, если смотреть на предметы, удаленные всего чем на десятки сантиметров.
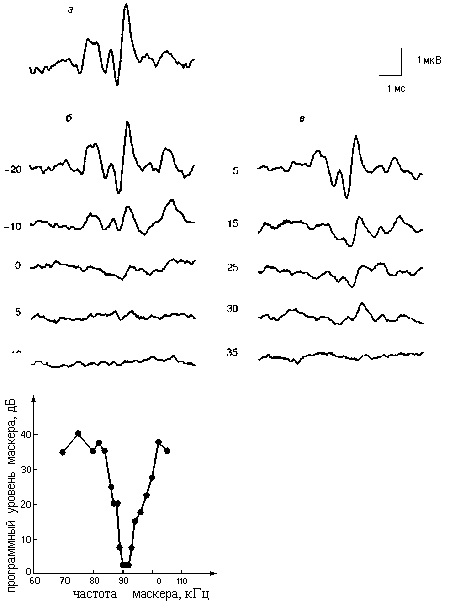
Итак, используя морфологическое исследование сетчатки, нам удалось не только оценить разрешающую способность зрения дельфинов, но и выявить необычную для млекопитающих организацию ОНВ и хотя бы отчасти понять, что представляет собой зрительный мир дельфина. Что различает слух дельфинаТо, что у дельфина необычайно развитый слух, известно уже десятки лет. Объемы тех отделов мозга, которые заведуют слуховыми функциями, у него в десятки(!) раз больше, чем у человека (при том, что общий объем мозга примерно одинаков). Дельфин способен воспринимать частоты звуковых колебаний, в 10 раз более высокие (до 150 кГц), чем человек (до 15-18 кГц), и слышит звуки, мощность которых в 10-30 раз ниже, чем у звуков, доступных слуху человека. Но чтобы ориентироваться в обстановке с помощью слуха, мало только слышать их, нужно еще тонко отличать один звук от другого. А эта способность дельфинов была исследована слабо, и мы постарались восполнить этот пробел. Напомним, звук - колебания воздуха, воды или другой среды. Любой естественный звук - это набор колебаний разных частот. От того, какими частотами колебаний он составлен, зависят его высота, тембр, т.е. то, чем один звук отличается от другого. Ухо животного или человека способно определять, из какого набора частот состоит звук: оно работает как набор частотных фильтров, каждый из которых откликается на свою частоту колебаний. Чтобы анализ был точным, настройка частотных фильтров должна быть острой. Чем острее настройка, тем меньшая разница частот доступна для различения, тем выше частотная разрешающая способность (ЧРС) слуха. Но звук - не просто набор разных частот. Каждая из них еще меняется во времени: становится то сильнее, то слабее. Слуховая система должна отслеживать эти быстрые изменения, и чем лучше она это делает, тем богаче сведения о свойствах звука. Поэтому помимо ЧРС, очень важна временная разрешающая способность (ВРС). Обе эти характеристики и определяют способность животного отличать один звук от другого. Именно эти характеристики слуха мы измеряли у дельфинов. Чтобы измерить какую-либо характеристику слуха, нужно решить две задачи: во-первых, узнать, слышит или не слышит животное пробный сигнал, а во-вторых, нужно подобрать пробные сигналы с такими свойствами, чтобы возможность услышать их зависела от измеряемого свойства слуха. Например, чтобы измерить чувствительность, нужно использовать звуки разной интенсивности: чем слабее звук, который удается услышать, тем чувствительность лучше. Чтобы узнать, что слышит дельфин, мы использовали регистрацию электрической активности мозга. Каждая нервная клетка продуцирует электрические потенциалы. При воздействии звука много клеток возбуждается одновременно, и продуцируемые ими потенциалы складываются в довольно мощный электрический сигнал, называемый вызванным потенциалом (ВП). ВП можно зарегистрировать, прикоснувшись контактным электродом к поверхности головы; такая процедура совершенно безвредна для животного. ВП - хороший показатель того, слышит ли дельфин звук. Если после подачи звука возник ВП, значит слуховая система реагирует на этот звук. Если величина ВП падает, звук воспринимается на пределе возможного. Если ВП нет - скорее всего звук не слышен. Для измерения ЧРС используется прием, называемый маскировкой [8]. Сначала дается пробный сигнал - звук определенной частоты, который вызывает электрический ответ мозга - ВП (рис.4, а). Затем к звуку добавляется другой звук - помеха, заглушающая пробный сигнал, который становится хуже слышимым, при этом амплитуда ВП падает (рис.4, б, в). Чем сильнее помеха, тем сильнее заглушение.
Рис.4. Измерение частотной селективности слуха дельфина методом маскировки: а - ВП, возникающий в ответ на тональную посылку с частотой 90 кГц; б - ВП, возникающий в ответ на такую же тональную посылку, но на фоне помехи с частотой также 90 кГц (цифры показывают уровень помехи в дБ относительно пробного сигнала); в - то же, при частоте помехи 85 кГц; г - настроечная кривая на частоте 90 кГц, полученная методом маскировки Маскировка используется для измерения ЧРС потому, что она зависит от частотно-избирательных свойств слуха. Сравним рисунки 4, б и в. В одном случае частоты пробного звука и помехи совпадали ( б), а в другом немного различались ( в). Обе серии записей похожи, но цифры показывают: если частоты пробы и помехи различаются ( в), то для маскировки нужна помеха намного сильнее, чем при совпадающих частотах ( б). Это и есть проявление частотной избирательности: слуховая система способна разделить частоты пробного сигнала и помехи, если они различаются. Чем острее частотная селективность, тем резче ослабевает маскировка при различии частот пробы и помехи. Нужно найти, как пороги маскировки зависят от разницы в частотах пробы и помехи.
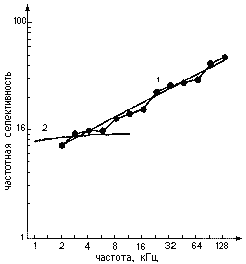
Рис.5. Зависимость частотной селективности от частоты звука у дельфина и у человека: 1 - частотная селективность звука дельфина (ломаная линия - экспериментальные данные, наклонная прямая - их аппроксимация); 2 - частотная селективность слуха человека (по литературным данным)
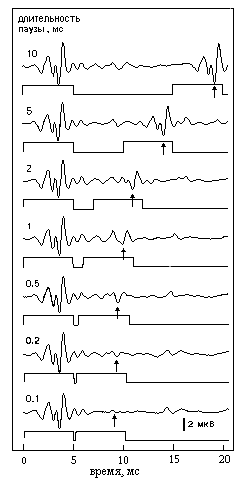
Результаты представлены в виде кривой (рис.4, г). Чем острее кривая, тем лучше частотная избирательность, выше ЧРС. А повторяя измерения с разными частотами пробы, можно измерить ЧРС во всем частотном диапазоне слуха (рис.5). При исследовании остроты слуховых фильтров, настроенных на разные звуковые частоты [9], прежде всего, была выявлена исключительно высокая ЧРС, особенно в области высоких звуковых частот (десятки кГц). Здесь уровень ЧРС доходит до 50 единиц, т.е. слух дельфина различает частоты, разнящиеся примерно на 1/50. Это в 4-5 раз лучше, чем у других животных и у человека. Но такая высокая избирательность наблюдается только в области высоких звуковых частот, не воспринимаемых человеком. В том диапазоне, который доступен и человеку, и дельфину, избирательность слуха дельфина заметно меньше - примерно как у человека. А как измерить временную разрешающую способность слуха? Для этого есть несколько способов. Можно использовать пары коротких звуковых импульсов: если интервал между импульсами в паре больше некоторой величины, то они слышатся раздельно, а если меньше, то сливаются в один щелчок. Тот минимальный интервал, при котором можно расслышать два отдельных импульса, - мера ВРС. Можно использовать звук, интенсивность которого ритмически пульсирует: та предельная частота пульсаций, при которой они еще не сливаются в монотонный звук, - тоже мера ВРС. Еще один способ: в непрерывном звуке делается короткая пауза. Если длительность паузы очень мала, то она "проскакивает" незамеченной. Минимальная длительность паузы, при которой ее можно обнаружить, - тоже мера ВРС. Мы использовали все эти способы. А чтобы узнать, слышит ли животное повторный звуковой импульс, или пульсации громкости, или короткую паузу, тоже регистрируются ВП мозга [10]. Например, чем короче пауза, тем ВП меньше, пока не исчезнет совсем (рис.6). Так же определяется слышимость и других пробных сигналов.
Рис.6. Слуховые ВП, возникающие у дельфина в ответ на короткую паузу в шуме. В каждой паре осциллограмм верхняя - регистрация биопотенциалов, нижняя - отметка включения-выключения звука (сдвиг вверх - включение, сдвиг вниз - выключение). Длительность паузы между двумя звуковыми посылками указана цифрами рядом с осциллограммами. Стрелки указывают ответы на паузу (они возникают с задержкой около 4 мс)
Мы получили впечатляющие результаты: ВРС у дельфина оказалась не в два-три и даже не в десять раз выше, чем у человека, а в десятки (почти в сто) раз. Слух человека позволяет различать интервалы времени примерно от одной сотой секунды (10 мс). Дельфины же различают интервалы в десятитысячные доли секунды (0.1-0.3 мс). Разрыв в звуке, длящийся 0.1 мс, вызывает хоть и небольшой, но различимый ВП. То же наблюдается и при действии других пробных звуков. Два коротких звуковых импульса отличаются от одного, когда интервал между ними составляет всего 0.2-0.3 мс (у человека - несколько мс). Пульсации громкости звука вызывают ответы, когда их частота приближается к 2 кГц (у человека - 50-70 Гц). Почему слуховая система дельфина имеет такие уникальные возможности? У человека и многих лабораторных животных ЧРС и ВРС слуха довольно близки, и создавалось впечатление, что это предел возможного для природы. Но слуховой системе дельфинов доступны и намного более острая частотная настройка, и лучшее различение интервалов времени. Почему же таких показателей не достигла слуховая система других животных? Видимо, все дело в неизбежном противоречии между частотной и временной разрешающей способностью: чем лучше ЧРС, тем хуже ВРС, и наоборот. Эта чисто математическая закономерность справедлива для любой колебательной системы: если система остро настроена на определенную частоту (высокая частотная селективность), то ее колебания медленно раскачиваются и затухают (низкое временное разрешение). Это можно выразить простым соотношением:
где Q - острота частотной селективности, F - частота, на которую настроен фильтр, и B - полоса пропускания фильтра, т. е. тот набор частот, который он пропускает. Темп, с которым может меняться амплитуда сигнала, зависит от полосы B: чем она шире, тем более быстрые изменения сигнала пропускаются фильтром, но тем "тупее" фильтр (меньше Q). Поэтому слуховая система должна найти некоторый компромисс между ЧРС и ВРС, ограничив обе эти характеристики на некотором уровне. Улучшение одной из них возможно только за счет ухудшения другой. Но противоречие между ЧРС и ВРС становится менее драматическим по мере увеличения частоты F: при высокой частоте можно сочетать широкую полосу B с острой избирательностью Q. Именно это и характерно для слуха дельфина, который освоил простор ультразвуковых частот. Значит, ЧРС и ВРС реально обусловлены не пределом возможного для слуховой системы, а разумным компромиссом между этими двумя характеристиками. Так исследование, казалось бы, экзотического животного позволяет понять фундаментальные принципы построения слуховой системы всех животных и человека. ЛИТЕРАТУРА
| ||||||||||
| World of Pern © is copyright Anne McCaffrey 1967, 2001. The Dragonriders of Pern ® is a registered trademark. | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|